


Fotosintesis adalah serangkaian reaksi biokimia yang digunakan autotrof fotosintesis untuk mengubah energi matahari menjadi energi potensial, dan itu melibatkan dua tahap.
Pada tahap pertama, yang disebut reaksi bergantung cahaya, autotrof menangkap energi dari sinar matahari. Dan pada tahap kedua, reaksi terang-independen atau gelap , energi matahari yang ditangkap diubah menjadi energi potensial, yang ada di autotrof sebagai ikatan kimia dalam makromolekul.
Sebagian besar makromolekul yang disintesis dari fotosintesis adalah karbohidrat, tetapi autotrof juga dapat menghasilkan protein dan lipid. Makromolekul ini dikonsumsi oleh autotrof dalam reaksi metabolisme mereka dan digunakan oleh heterotrof sebagai makanan dan produk non-makanan lainnya.
| Glosarium | |
|---|---|
Kloroplas | Sebuah organel membran ganda pada eukariota fotosintesis seperti tanaman di mana fotosintesis berlangsung. |
Granum (jamak grana) | Tumpukan struktur seperti kantung pipih yang disebut tilakoid di dalam kloroplas. |
Proplastida | Organel yang belum matang ditemukan pada autotrof eukariotik yang tidak mengandung pigmen, yang matang dan berdiferensiasi menjadi kloroplas dan plastida lainnya. |
foton | Partikel elementer atau kuantum cahaya yang menampilkan sifat-sifat gelombang dan partikel. Ini dianggap sebagai jumlah cahaya terkecil. |
Fosforilasi | Proses kimia pengikatan gugus fosfat dalam sel. Kebalikan dari reaksi ini disebut defosforilasi. |
Fotoreseptor | Kompleks protein peka cahaya yang merasakan dan merespon keberadaan cahaya. |
Fotofosforilasi | Reaksi kimia seluler yang mensintesis ATP dari fosforilasi ADP menggunakan energi matahari. |
Tilakoid | Struktur berbentuk cakram berongga yang terikat membran di kloroplas tempat fotosintesis pada eukariota berlangsung. |
Stroma | Cairan yang mengisi bagian dalam membran dalam kloroplas. |
Apa itu Fotosintesis?
Fotosintesis terjadi pada autotrof , yaitu organisme yang mampu menghasilkan makanannya sendiri. Mereka termasuk archaea, prokariota seperti bakteri ungu, acidobacteria, dan cyanobacteria, dan eukariota seperti ganggang dan tanaman darat.
Fotosintesis dapat berupa anoksigenik atau oksigenik. Fotosintesis anoksigenik tidak memerlukan air atau menghasilkan O2 , dan selain cyanobacteria, fotosintesis pada non-eukariota bersifat anoksigenik .
Fotosintesis oksigenik dianggap sebagai pembalikan glikolisis dalam-dalam respirasi seluler. Ini menangkap energi dari sinar matahari dan menggunakannya untuk mengubah CO 2 menjadi gula dan oksigen dengan adanya air.
Ada dua tahap fotosintesis, yang dapat diringkas sebagai berikut: [1,2]
(CO) 2 + nH 2 O →(CH 2 O) n +O 2 ;
dimana n menyatakan jumlah molekul.
Pada tahap pertama ( tergantung cahaya) , fotoreseptor memanen energi dari sinar matahari, dan energi yang dipanen mengoksidasi air menjadi oksigen (O 2 ) dan memicu transfer elektron. Hal ini menyebabkan reduksi n icotinamide a denine d inucleotide p hosphate (NADP + ) dan sintesis denosin t riphos p benci (ATP) dari fosforilasi denosin d iphos p benci (ADP) , dalam reaksi yang disebut fotofosforilasi.
Tahap kedua, reaksi tergantung cahaya , adalah bagian dari siklus Calvin . Reaksi terjadi tanpa keterlibatan langsung sinar matahari. Sebaliknya, ATP dan NADPH yang dihasilkan dalam reaksi terang dikonsumsi untuk mensintesis gula dari karbon dioksida (CO 2 ). ADP dan NADP + yang dihasilkan didaur ulang untuk memasok transfer elektron dalam reaksi yang bergantung pada cahaya. [1]
Fotosintesis pada Eukariota Terjadi di Kloroplas
Fotosintesis pada eukariota terjadi di kloroplas, organel yang secara genetik mirip dengan cyanobacteria, yang mengarah ke hipotesis endosimbiotik yang mengatakan bahwa asal usul kloroplas sebanding dengan mitokondria.
Secara khusus, kloroplas berasal dari eukariota heterotrofik yang mengambil cyanobacteria fotosintesis. Setelah endosimbiosis, bakteri kehilangan kemampuan untuk hidup mandiri, mengubah eukariota inang dari heterotrof menjadi autotrof . [3]

Gambar 1: Fitur Struktural Kloroplas pada Autotrof Fotosintetik
Mirip dengan mitokondria, kloroplas memiliki dua lapisan membran, membran luar dan dalam. Membran luar membungkus stroma , yang mengandung beberapa grana dan protein lainnya. Setiap granum terdiri dari tumpukan tilakoid , struktur seperti kantung pipih, dan ruang berair di setiap tilakoid disebut lumen tilakoid ; namun, lingkungan di dalam lumen tilakoid bersifat asam (Gambar 1) .
Kloroplas hadir dalam sel dan jaringan yang terlibat dalam fotosintesis, misalnya, sel penjaga dan mesofil di epidermis daun. Mereka dibedakan dari plastida tidak berwarna yang belum matang, yang disebut proplastida, dan mengandung pigmen yang didominasi klorofil , yang memberi warna hijau pada tumbuhan. [1,2]
Pigmen fotosintesis menangkap foton.
Matahari memancarkan energi sebagai gelombang elektromagnetik, terdiri dari beberapa panjang gelombang yang berbanding terbalik dengan tingkat energi. Unit energi terkecil yang dibawa oleh cahaya dikuantifikasi dalam bentuk kuanta cahaya atau foton . Fotosintesis di sebagian besar autotrof menggunakan cahaya tampak, yang panjang gelombangnya berkisar antara 350 hingga 800 nanometer. [4]
Klorofil berfungsi sebagai pigmen utama yang menyerap cahaya dan menangkap foton dalam alga, cyanobacteria, dan tumbuhan. Mereka memiliki cincin porfirin yang terdiri dari empat pirol yang berkoordinasi dengan satu ion magnesium (Mg 2+ ) di tengahnya.
Cincin tersebut memiliki rantai samping hidrokarbon yang panjang dengan satu ikatan karbon-karbon ganda yang memungkinkan klorofil dilarutkan dalam lipid dan tertanam dalam membran tilakoid. [1]
Dua spesies klorofil yang dominan ditemukan pada tumbuhan dan alga: [1]
- Klorofil a dibedakan oleh gugus metil yang melekat pada salah satu dari empat cincin pirol. Ini adalah spesies klorofil yang paling umum pada tumbuhan dan satu-satunya yang ada di pusat reaksi fotosintesis . Klorofil a menyerap cahaya paling banyak pada 372 dan 642 nanometer, masing-masing sesuai dengan cahaya ungu-biru dan oranye. [1,4]
- Klorofil b dicirikan oleh gugus formil yang melekat pada cincin porfirin pada posisi yang sama dengan gugus metil pada klorofil a. Substitusi gugus formil menggeser rentang penyerapan cahaya optimal dalam klorofil b menjadi 392 dan 626 nanometer, masing-masing sesuai dengan lampu biru dan merah. [1,4]
Selain klorofil , pigmen aksesori lain juga ada dalam sel fotosintesis. Pigmen aksesori ini menyerap cahaya tampak dari panjang gelombang yang berbeda, mendukung proses penangkapan energi. Pigmen aksesori dapat mencegah cahaya yang intens atau berlebihan dari kerusakan klorofil dan komponen fotosintesis dengan mengurangi radikal bebas. [1, 2]
Contoh pigmen aksesori adalah: [1,2]
- Karotenoid adalah pigmen yang menyerap cahaya biru-hijau dan ungu, sesuai dengan 400 hingga 500 nanometer. Mereka memberi ganggang dan tanaman warna kuning, oranye, dan merah. Contohnya adalah karoten yang merupakan karotenoid hidrokarbon tak jenuh, dan xantofil yang merupakan karotenoid teroksigenasi.
- Phycobilins adalah pigmen merah yang menyerap cahaya dalam kisaran 550 hingga 630 nanometer. Tidak seperti karotenoid, fikobilin hanya ditemukan pada alga merah dan beberapa alga uniseluler (seperti cryptomonads) tetapi tidak pada tumbuhan atau alga hijau.
Karena semua pigmen fotosintesis menyerap cahaya pada panjang gelombang yang berbeda, aksi penyerapan cahaya gabungan dari semua pigmen akan memungkinkan organisme untuk memaksimalkan penangkapan foton. [1]
Mekanisme Fotosintesis
Mekanisme fotosintesis dapat dibagi menjadi dua tahap berdasarkan kebutuhan cahaya. Kedua tahap fotosintesis dihubungkan oleh dua molekul berenergi tinggi, ATP dan NADPH (Gambar 2) . Baik ATP dan NADPH dikonsumsi pada tahap kedua ketika gula heksosa disintesis. Gula heksosa adalah blok bangunan untuk sintesis karbohidrat kompleks .
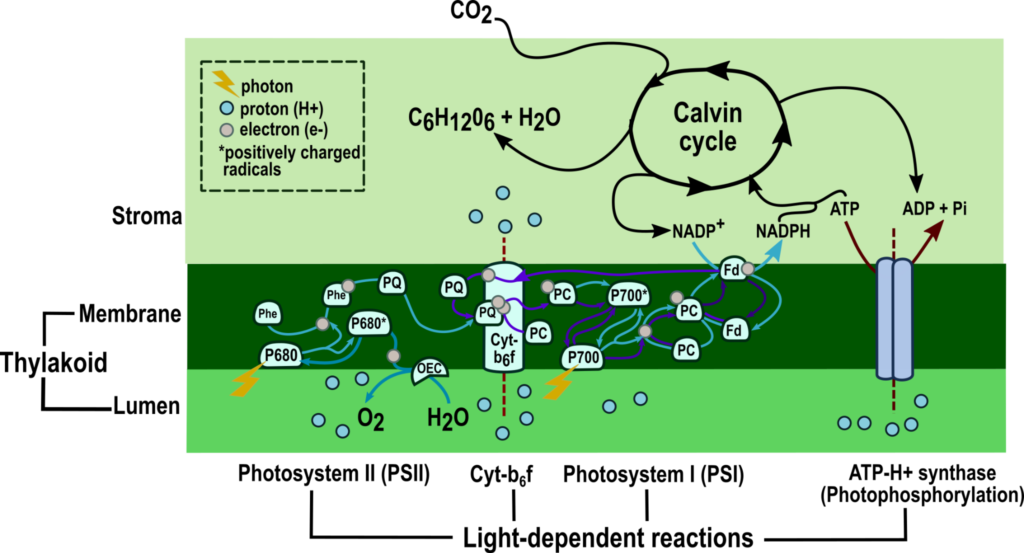
Gambar 2: Mekanisme fotosintesis — Ringkasan dua tahapnya: reaksi bergantung cahaya dan siklus Calvin. Tahap pertama terdiri dari pemanenan cahaya, transfer elektron dan fotofosforilasi. Foton diambil dari cahaya oleh fotoreseptor di fotosistem I dan II, memicu transfer elektron antara dua fotosistem dan kompleks protein, sitokrom b 6 f . Transfer elektron menghasilkan gradien proton, yang mendorong fotofosforilasi, seperti yang digambarkan oleh panah merah tua. Fotofosforilasi non-siklik, yang dikenal sebagai skema Z, dihasilkan dari transfer elektron yang ditunjukkan oleh panah biru muda, dan fotofosforilasi siklik dihasilkan dari transfer elektron yang diwakili oleh panah ungu. Reaksi terang menghasilkan ATP dan NADPH yang dikonsumsi dalam siklus Calvin ketika gula heksosa (C 6 H 12 O 6 ) dihasilkan.
A. Reaksi Tergantung Cahaya
Seperti yang ditunjukkan dalam namanya, reaksi fotosintesis bergantung pada cahaya terjadi ketika ada cahaya. Pada eukariota, reaksi bergantung cahaya berlangsung di membran tilakoid dalam tiga langkah (Gambar 2) : [1]
1. Memanen Foton dari Cahaya
2. Transfer Elektron
- PSI (Plastosianin-feredoksin oksidoreduktase)
- PSII (Air-plastoquinine oksidoreduktase)
- Kompleks sitokrom b6f (Plastoquinone-plastocyanin oxidoreductase)
PSI (Plastocyanin-Ferredoxin Oxidoreductase)
Setelah itu, PC teroksidasi menyumbangkan elektron ke akseptor berikutnya yang tersedia. Setelah elektron mencapai sisi stroma membran tilakoid, elektron mengoksidasi ferredoxin (Fd), sementara NADP + di stroma secara bersamaan direduksi menjadi NADPH.
NADPH yang diproduksi di PSI selanjutnya digunakan dalam sintesis karbohidrat. P700 yang bermuatan positif disuplai kembali dengan elektron yang ditransfer dari Cyt-b 6 f .
PSI (Plastocyanin-Ferredoxin Oxidoreductase)
Kompleks sitokrom b6f (Plastoquinone-Plastocyanin Oxidoreductase)
3. Fotofosforilasi[1, 2]
Jalur skema Z (fotofosforilasi non-siklik)
dimana n menyatakan jumlah molekul.
Fotofosforilasi siklik
B. Siklus Calvin(-Benson-Bassham)
1. Asimilasi Karbon Dioksida
2. Sintesis Pentosa Reduktif
3. Regenerasi Molekul Akseptor Karbon
Fotorespirasi adalah Calvin Cycle Bad Company
Faktor-Faktor yang Mempengaruhi Fotosintesis
Laju fotosintesis tergantung pada efisiensi dan efektivitas kedua tahapnya. Berdasarkan mekanismenya, beberapa faktor lingkungan dan internal mempengaruhi dua tahap fotosintesis, antara lain:
1. Kualitas dan Kuantitas Cahaya
2. Ketersediaan Air
3. Konsentrasi Karbon Dioksida
4. Suhu
5. Predisposisi Genetik
Kesimpulannya
Referensi:
- Boyer R, Konsep dalam Biokimia, edisi ke-3. New Jersey: John Wiley & Sons; 2006.
- Dimiliki H-Wa. Biokimia Tumbuhan. edisi ke-3 San Diago, California: Pers Akademik; 2005.
- Martin WF, Garg S, Zimorski V. Teori endosimbiotik untuk asal eukariota. Philos Trans R Soc B Biol Sci. 2015;370(1678):20140330. doi:10.1098/rstb.2014.0330
- Milne BF, Toker Y, Rubio A, Nielsen SB. Mengungkap Warna Intrinsik Klorofil. Angew Chemie Int Ed. 2015;54(7):2170-2173. doi:10.1002/anie.201410899
- Gao J, Wang H, Yuan Q, Feng Y. Struktur dan Fungsi Fotosistem Superkompleks. Ilmu Tanaman Depan. 2018;9. doi:10.3389/fpls.2018.00357